 "
"
Team:CIDEB-UANL Mexico/math union
From 2014hs.igem.org
|
|








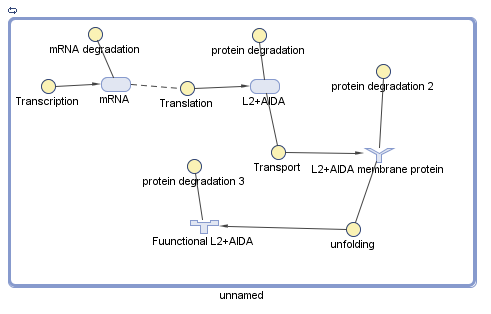
Union Module
The union module is based on the use of a fusion protein composed by L2 and AIDA. L2+AIDA is a protein which is not affected by external factors during its transcription, as well as its translation, so the established parameters was used with the data obtained by the team:
\begin{equation} \large \frac{d\left [ mRNA \right ]}{dt}= \alpha_{1}-d_{1}\left [ mRNA \right ] \end{equation}
The parameters for both, the translation and transcription rate from Singapore 2008 iGEM team, were used, as well as the speeds at which E. coli carries out the transcription and translation, assuming a transcription speed of 70nt/s and a translation speed of 40aa/s. These data were used in the equations below with the L2+AIDA gene length of 2620nt and protein length of 856aa:
\begin{equation}
\large \alpha_{1} = \frac{transcription speed}{gene length (nt)}
\end{equation}
\begin{equation}
\large \alpha_{2} = \frac{translation speed}{protein length (aa)}
\end{equation}
\begin{equation}
\large \alpha_{1} = \frac{(70)(60)}{2620} = 1.6
\end{equation}
\begin{equation}
\large \alpha_{2} = \frac{(40)(60)}{856} = 2.8
\end{equation}
Then, the parameters for degradation rates for proteins and mRNAs from Beijing PKU 2009 iGEM team were used:
\begin{equation} \large d_{1} = \frac{1}{half-life(min)} + \frac{1}{30min} \end{equation}
\begin{equation} \large d_{2} = \frac{1}{half-life(min)} + \frac{1}{30min} \end{equation}
Since the protein was the fusion of two, a research was needed to find out the half-life each protein separately. The half-life of membrane proteins range between 2 to 20 hours in E. coli (Hare, 1991), and as AIDA-I is a membrane protein its half-life must be between that range, for the specific half-life of AIDA is not determined. To find the half-life of L2, based on previous research, it was assumed that it was 7.8 hours (Bergant, 2010). Bergant’s team made a test with a homologous protein, but found in the minor capsid of the Human Papillomavirus (HPV). Although the function of the L2 strand in HPV is viral and in E. coli is ribosomal, both share similar structures and sequences. Once it was decided to use the half-life from the homologous L2, it was used as the half-life for the fusion protein because it was between the range of AIDA-I and also because it was the lower half-life, assuming E. coli starts the degradation of L2, the whole protein would be degraded.
To determine the degradation rate of average mRNA, the information from Selinger’s team (2003) was used. They carried several experiments to find out the average mRNA half-life in E. coli. They used mRNAs about 1100nt, concluding they have an average half-life of 5min. So with this, the average mRNA half-life of L2+AIDA was found to be 11.9min.
\begin{equation} \large HL = \frac{1100(nt)}{5 min} \end{equation}
With all these information, the degradation rates for both transcription and translation of L2+AIDA were found:
\begin{equation}
\large d_{1} = \frac{1}{11.9} + \frac{1}{30} = 0.11
\end{equation}
\begin{equation}
\large d_{2} = \frac{1}{468} + \frac{1}{30} = 0.035
\end{equation}
Simbiology was used for the simulation. The previous data from the equations were used to calculate the amount of proteins E. coli would produce at certain times. The results obtained are shown in the following graph:
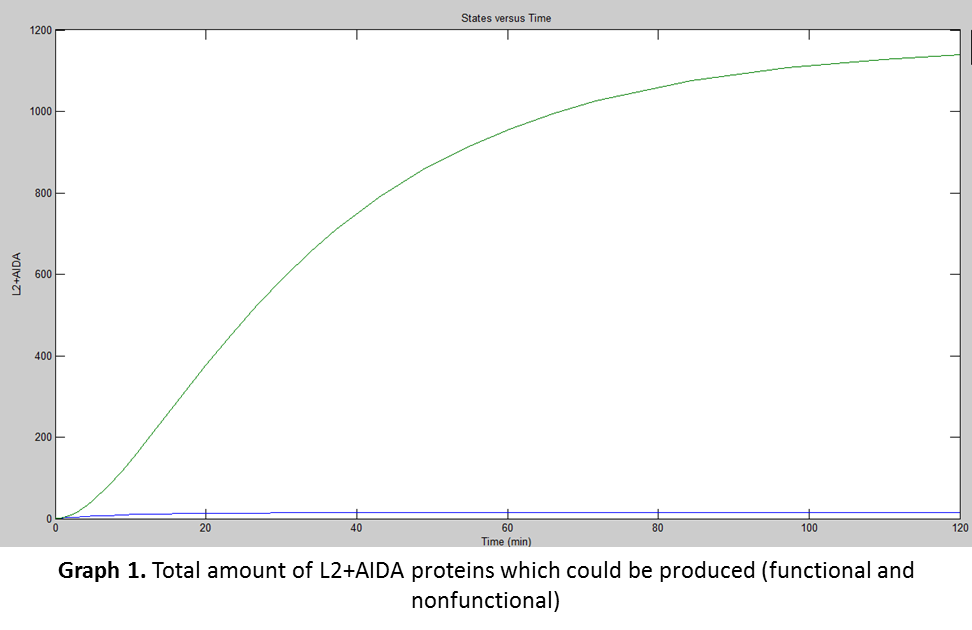
For translation there was another factor that was needed to be taken into consideration, the “fpost”, which were the posttranslational variables affecting the production of the functional protein:
\begin{equation}
\large \frac{d[P]}{dt} = \alpha_{2} \cdot[mRNA] - d_{2}[P] - f_{post}
\end{equation}
Since the fusion protein needs to be expressed in the membrane of E. coli, the average velocity at which E. coli exports its proteins to the membrane was needed. The process by which bacteria exports its proteins are divided into three phases: the “breathing” between translation and the second phase, which is movement of a protein to the membrane, and translocation (in this phase the protein attaches to the membrane of the bacteria (Peskin, 1991)). Using this information, it was determined that E. coli completes these phases in an average of 5 to 6 min, depending on the protein size (Driessen, 1990). The time used was 5.5min because L2+AIDA are not too small or too big in length (2620nt).
According to Ikeda y Kuroda (2011), L2 carries out an unfolding process to become functional. It was found out that the 50S ribosomal proteins L2, L3, L14, L23, L24, and L32, as well as the 30S ribosomal proteins, S12 and S18, were native premolten globules in their free forms, but adopted rigid well-folded conformations during the formation of a functional ribosome. They exhibit several amounts of ordered secondary structures; the unfolding of a protein molecule results in an essential increase in its hydrodynamic volume. For instance, there is a well-documented 15–20% increase in the hydrodynamic radius of globular proteins upon their transformation into the molten globule state (Unversky, 2002). Also, the equation from Unversky to find the L2 unfolding rate in E. coli was used, with the results shown below:
\begin{equation}
\large [H]boundary = \frac{[R]+1.51}{2.785}
\end{equation}
This equation gives the estimation of the "boundary" mean hydrophobicity value, “[H]boundary”, below which a polypeptide chain with a given net charge “[R]” will most probably be unfolded. Thus, sequences of natively unfolded proteins may be characterized by a low sequence complexity and/or high net charge coupled with low mean hydrophobicity (the values are specified for globular proteins). According to Ikeda and Kuroda (2011) the net charge “[R]” of L2 is 10.9, so we substituted it in the equation below:
\begin{equation} \large [H]boundary = \frac{10.9 + 1.51}{2.785} = 4.45 \end{equation}
With the unfolding value and the rate of membrane transport in E. coli we could use it in Simbiology for modelling the functional L2+AIDA production. The results obtained were the following:

Comparing both graphs (Graph 1 and Graph 2) we realize that although E. coli needs to transport the L2+AIDA proteins to its membrane, the rate at which E. coli does it is slower than the production of the fusion protein, but something that was noticed (and was great) is that according to Simbiology, almost all the proteins, once they are inserted in the membrane, unfold correctly leaving less than 25 nonfunctional proteins.
Bibliography/References
● BERGANT, Martina N. M. (2010). Modification of Human Papillomavirus Minor Capsid Protein L2 by Sumoylation. Journal of Virology, 11585-11589.
● DRIESSEN, Arnold W. W. (1990). Proton transfer rate-limiting for translocation of precursor proteins by the Escherichia coli translocase. Biochemistry, 2471-2475.
● HARE, James K. T. (1991). Mechanisms of plasma membrane protein degradation: Recycling proteins are degraded more rapidly than those confined to the cell surface. PNAS, 5902-5906.
● IKEDA, Takeshi A. K. (2011). Why does the silica-binding protein "Si-tag" bind strongly to silica surfaces? Implications of conformational adaptation of the intrinsically disordered polypeptide to soli surfaces. Colloids and Surfaces, 359-363.
● PESKIN, Charles S. S. (1991). What drives the translocation of proteins. Biophysics, 3770-3774.
● SELINGER, Douglas R. M. (2003). Global RNA Half-Life Analysis in Escherichia coli Reveals Positional Patterns of Transcript Degradation. Genome Research, 216-223.
● UVERSKY, V. (2002). Natively unfolded proteins: A point where biology waits for physics. Protein Science, 739-756.
Av. Lázaro Cárdenas to the
East n/a Col. Mederos, Monterrey, Nuevo León, México.
Pedro de Alba n/a Ciudad Universitaria, San Nicolás de Los Garza, Nuevo León, México.

igem.cidebuanl@gmail.com




iGEM CIDEB UANL 2014. CENTRO DE INVESTIGACIÓN Y DESARROLLO DE EDUCACIÓN BILINGÜE. UANL